

Mechanisms of Developmental Biology Series: Epagoge in Development: From Embryology to Evolution
- Gülce Tekin

- Feb 21, 2024
- 18 min read
Updated: Sep 11, 2024
Foreword
The progression from a single-cell entity to a multicellular organism entails a series of intricate stages. This journey involves the orchestrated activities of stem cells and the process of differentiation, which collectively constitute the life cycle of complex multicellular organisms. The domain of developmental biology transcends human boundaries, encompassing studies of animals and plants alike, underscoring its extensive purview. The pursuit of unraveling the intricate mechanisms governing organismal development not only illuminates the origins of life but also provides invaluable insights into the molecular and cellular underpinnings of development across diverse species. The curriculum of this developmental biology series serves as an illuminating beacon, shedding light on the myriad manifestations of life on our planet. To gain a more profound comprehension of developmental processes, this series will traverse multiple interdisciplinary avenues, including embryology, medical science, and evolutionary biology. Through this holistic approach, we endeavor to uncover the underlying principles governing the diverse tapestry of life forms on Earth.
This series will be divided into the following sections:
5. Mechanisms of Developmental Biology Series: Epagoge in Development: From Embryology to Evolution
6. Mechanisms of Developmental Biology Series: Cutting-edge Techniques in Developmental Biology
Mechanisms of Developmental Biology Series:
Epagoge in Development: From Embryology to Evolution
Evolutionary developmental biology stands at the intersection of genetics, embryology, and evolution, unraveling the mechanisms governing the diversity of life forms. This comprehensive review explores key concepts, from the historical roots of embryology and evolution to the modern insights provided by genetic studies. The discovery of master regulatory genes, such as Hox genes, and the role of epigenetic processes have revolutionized our understanding of how genetic changes translate into morphological variations. The link between development and evolution, reinterpreted through evo-devo principles, offers a nuanced perspective on homology and shared ancestry. This interdisciplinary approach provides a holistic framework for comprehending the unity and adaptability observed in living organisms, emphasizing the dynamic interplay between genetic foundations and environmental influences.

The Idea of Evolutionary Developmental Biology
Evolutionary developmental biology (evo–devo) involves the comparative examination of the development of various organisms and their changes throughout evolution, by specifically emphasizing the genetic foundation of phenotypic structures, exploring their transformations during evolutionary processes and the emergence of new structures. All of the transformations in the form and function of animals, throughout evolution, stem from alterations in their DNA. These genetic changes dictate the production, timing, and location of proteins during embryonic development, exerting control over the behavior of embryonic cells. In the evolutionary process, modifications to organs typically entail adjustments to the development of pre-existing structures, essentially tinkering with what already exists. A prime illustration of this phenomenon is evident in the evolution of jaws, which originated from the pharyngeal arches of jawless ancestors, and the transformation of the incus and stapes in the middle ear from bones originally situated at the joint between upper and lower jaws (Wolpert, 2000).
The link between evolution and development is in fact partly linguistic: the term "evolution," (derived from the Latin words "evo/utio" meaning "unrolling") was initially employed to describe the emergence of an individual organism during embryonic development. Early theories that emphasized minimal interaction with the external environment and prioritized constancy and destiny applied this term attributed to a Swiss scientist, Albrecht von Haller (1708–1777), who introduced the term "evolution" in 1774 to characterize the development of the individual within the egg (Hall, 1999).

History of Evo-Devo
A new era began in the scientific circles in England with the publication of Charles Darwin's The Origin of Species in 1859. In in 1866, Ernst Haeckel came up with a theory that puts forward the developmental stages of an organism during its embryonic and fetal development (ontogeny). He claimed that these stages mirror or recapitulate the evolutionary stages of its species (phylogeny), which formed the foundation of his Biogenetic Law. The first empirical test of Darwin's theory, using embryos and larvae, conducted by Fritz Müller in 1864. It revealed that ontogeny could provide insights into phylogeny. As opposed to Haeckel, Müller's study demonstrated the potential to understand evolutionary patterns and identify mechanisms through ontogenetic exploration. Starting from the mid-20th century, the fusion of Mendelian and population genetics seemed to provide explanations for maintaining organismal features, variations in those features, and the origin of new features.
Stephen J. Gould's book, Ontogeny and Phylogeny, published in 1977, highlighted the concept of heterochrony which was first established by Haeckel (Hall, 2012). This concept argues changes in the timing of developmental processes between the descendants and ancestors of a particular lineage, such as certain features appearing earlier or later in the development of an organism compared to its ancestors, influencing its overall morphology. This concept can be considered an important evolutionary developmental mechanism, demonstrating how alterations in the timing of developmental processes can contribute to evolutionary change. An example of heterochrony could be axolotls, which the juvenile external gills persist into adulthood and serve various functions including reproduction although most salamanders lose these gills when they metamorphose into adults.

A major development in evo-devo emerged when it was discovered that all animals (later extended to plants and fungi) share genes containing a 180-bp sequence, which was discovered in the fruit fly Drosophila. These genes, referred to as homeobox, homeotic, or Hox genes, play a crucial role in determining key body axis features, such as anterior and posterior sides, and specific body regions . They are pivotal in determining cell identity and positioning of body structures throughout embryonic development. Mutations in these genes have the potential to induce significant developmental abnormalities, leading to the loss of specific structures or alterations in the identity of body parts or segments commonly referred to as "homeotic transformations" (Duverger & Morasso, 2008).
The discovery of master regulatory genes—genes that should not be under the regulatory influence of any other gene (Chan & Kyba, 2013)—also contributed to the understanding of genetic conservation across animals whose morphology vary enormously (Hall, 2012). Over the last two decades, the key objective and challenge for evo-devo have been to combine the advancing molecular knowledge with the mechanisms that operate at various levels in order to build a bridge between genotype and phenotype.
Epigenetics
The idea of epigenetics has played a vital role in evo-devo, especially before the advanced molecular genetic revolution in developmental biology. The term "epigenetic" means "in addition to changes in genetic sequence", referring to any process that modifies gene activity without altering the DNA sequence which may lead to heritable modifications in daughter cells. There remains an ongoing debate about the precise definition and the scope of the term since some epigenetic modifications can be reversed (Weinhold, 2006). The reason for why not all of our cells and tissues are not identical relies on the expression pattern of genes. Epigenetic silencing serves as a mechanism to deactivate genes, contributing to this variation in gene expression.

Numerous types of epigenetic processes exist, including methylation, acetylation, phosphorylation, ubiquitylation, and s-acylation. While epigenetic processes are natural and crucial for many organism functions, improper occurrences can result in significant adverse health and behavioral effects. Within cells, there are three systems that can interact with each other to silence genes: DNA methylation, histone modifications, and RNA-associated silencing. Epigenetic modifications frequently occur throughout an organism's lifetime, and when these alterations take place in germ cells, they can be inherited by the next generation. Epigenetic processes, including "paramutation, bookmarking, imprinting, gene silencing, X chromosome inactivation, position effect, changeable disorder or phenotypic severity, reprogramming, maternal attributes, carcinogenic processes, teratogenic effects, regulation of histone modifications, heterochromatin states, and cloning" are recognized for their involvement in various biological phenomena (Moosavi & Motevalizadeh, 2016).
DNA Methylation
DNA methylation, confirmed to play a role in cancer, represents adding or removing a methyl group (CH3), particularly where consecutive cytosine bases occur. Methylation is catalyzed by DNA methyltransferase enzymes, involving the addition of a methyl group directly to a cytosine nucleotide within a cytosine-guanine sequence (CpG), often surrounded by other CpG’s forming a CpG island (Al et al., 2019). The presence of a methylated cytosine can repress transcription by inhibiting the binding of transcription factors or may promote the binding of other transcriptional repressors, including histone-modifying proteins (Fedoriw et al., 2012). Enzymes involved in the processes of establishing, recognizing, and removing DNA methylation can be categorized into three classes: writers, erasers, and readers. Writers refer to enzymes that facilitate the addition of methyl groups onto cytosine residues, erasers are responsible for modifying and eliminating the methyl group, and readers are enzymes that recognize and bind to methyl groups, ultimately influencing gene expression (Moore et al., 2012). The covalent addition of a methyl group to the C5 position of cytosine usually takes place in regions containing CpG dinucleotides, especially within regulatory sequences that play a role in suppressing gene expression. CpG methylation is crucial for repressing the transcription of transposons and repeat elements and for processes such as autosomal imprinting, X-chromosome inactivation, and the regulation of tissue-specific gene expression during developmental and differentiation stages (Loscalzo & Handy, 2014).

Utilizing mouse as a model system to investigate the role of DNA methylation in mammals, research has indicated that primordial germ cells, embryonic stem cells, and blastocyst cells appear to undergo normal development without detectable DNA methylation. However, as soon as stem cells initiate the differentiation process, DNA methylation becomes crucial for proper development. Many repetitive elements exhibit consistent DNA methylation patterns that persist throughout embryonic development without significant alterations. Likewise, imprinted genes manage to bypass the initial round of epigenetic reprogramming by preserving their epigenetic marks (Paulsen & Ferguson-Smith, 2001). Imprinting is defined as the epigenetic marking of the parental genomes of a diploid organism concerning their parental origin. This phenomenon influences the expression of certain genes in an organism, with imprinted genes being preferentially expressed from only one of the parental copies. DNA methylation patterns are present in almost all imprinted gene clusters and play a pivotal role in defining parental identity (Elhamamsy, 2017).

Histone Modifications
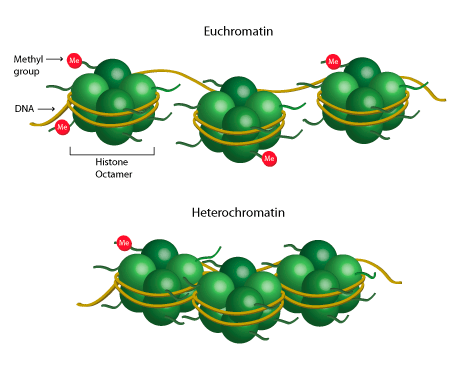
Chromatin is composed of a complex of nuclear DNA and histone protein, where DNA is compact and highly compressed by wrapping around histone proteins. The chromatin structure is not static but varies dynamically, even within interphase chromosomes where the cell is not actively dividing. It consists of two main states of chromatin: heterochromatin and euchromatin (Kitazawa & Kitazawa, 2023). Heterochromatin represents tightly compacted chromatin regions that are highly condensed and appear dark under a microscope. Heterochromatin is associated with transcriptional repression, meaning that genes within these regions are less accessible for transcriptional machinery, leading to lower gene expression. Euchromatin, in contrast, is a more open and accessible form of chromatin (less condensed and appears lighter under a microscope). It is associated with active transcription and high gene expression (Kouzarides, 2007).
Histones undergo various post-translational modifications, such as acetylation, methylation, phosphorylation, and ubiquitination, resulting in changes to chromatin structure that impact gene expression. For example, histone acetylation commonly takes place at lysine residues that carry a positive charge, resulting in the weakening of interactions between DNA and histones, ultimately leading to the opening of chromatin and facilitating transcription (Bannister& Kouzarides, 2011). Likewise, methylation of histones can take place at different sites within histone proteins, predominantly targeting lysine and arginine residues. This process is intricately regulated by numerous positive and negative factors, acting even at a specific site, with the ability to either activate or repress transcription (Jambhekar et al., 2019). So, chemical modifications to these histones result in the conversion of DNA from its euchromatin state to its heterochromatin state. The "Histone Code" hypothesis proposes that different types and combinations of modifications selectively modify chromatin structure and transcriptional potential (Loscalzo & Handy, 2014). By employing these mechanisms, alterations in a person's phenotype can occur without any changes to their fundamental genetic composition, effectively regulating gene expression (Shahid et al., 2024).
RNA-associated Silencing
RNA silencing is a sophisticated gene regulatory mechanism that operates at both transcriptional and posttranscriptional levels, namely Transcriptional Gene Silencing (TGS) and Post-transcriptional Gene Silencing (PTGS), also known as RNA interference (RNAi) (Agrawal et al., 2003). The RNA interference (RNAi) pathway can be triggered by various endogenous signals, including foreign DNA, double-stranded RNA (dsRNA) from viruses, abnormal transcripts from repetitive genomic sequences like transposons, and precursor forms of microRNA (pre-miRNA). In plants, RNAi is integral to virus-induced gene silencing (VIGS), playing a crucial role in defending against pathogens. Therefore, the presence of a foreign DNA is essential for RNAi mechanisms to be triggered. Studies on Caenorhabditis elegans suggest a potential mechanism for RNAi-mediated regulation of endogenous genes.
A simplified model of the RNAi pathway involves two main steps, both facilitated by ribonuclease enzymes that cause the degradation of RNA into smaller pieces. In the first step, the initial RNA trigger, then either dsRNA or the primary transcript of a microRNA (miRNA) undergoes processing by the RNase III enzymes Dicer and Drosha. This processing results in the generation of short interfering RNA (siRNA) that integrate into the RNA-induced silencing complex (RISC), where the siRNA unwinds. The single-stranded RNA from siRNA then pairs with its target mRNA. The gene silencing effect occurs through the degradation of the targeted mRNA, which is catalyzed by the enzyme Argonaute, also known as Slicer. Mismatches between the siRNA and mRNA duplex would cause cleavage not to occur (NCBI, 2017).

During interactions between plants and pathogens, plants must defend themselves against living transposable elements targeted by these pathogens. To counteract the aggressiveness of biotic stressors, plants employ various defense mechanisms when responding to these elements. RNA interference (RNAi) stands out as a crucial biological process in plants, capable of inhibiting gene expression at both transcriptional and post-transcriptional levels. This defense mechanism involves distinct groups of proteins that play a pivotal role in resisting the virulence of pathogens .However, pathogens have developed counteractive strategies to thwart this plant defense mechanism. They activate an anti-silencing mechanism by expressing suppressors, effectively blocking the host's RNAi. The disruption of the silencing mechanism serves as a virulence strategy employed by pathogens to enhance infection in the invaded hosts (Muhammad et al., 2019).
In Arabidopsis and other plant species, a variety of AGOs, DCLs, and RDRs have been identified (Baulcombe, 2004), suggesting the existence of specialized RNA silencing pathways, particularly for cellular movement. Presently, compelling evidence supports the existence of at least four distinct RNA silencing pathways in plants, each associated with different types of small RNAs, namely microRNA (miRNA) and small interfering RNA (siRNA) (Melnyk et al., 2011).
X-chromosome Inactivation
In contrast to the gene-scarce Y chromosome, the X chromosome boasts over 1,000 genes crucial for proper development and cell viability. Nevertheless, females possess two copies of the X chromosome, leading to a potential overdose of X-linked genes. To address this imbalance, female mammals have developed a distinctive mechanism for dosage compensation, diverging from strategies employed by organisms like flies and worms, through a process known as X-chromosome inactivation (XCI). Here, female mammals coordinately silence one of their two X chromosomes and the inactivated X chromosome subsequently undergoes condensation, forming a compact structure referred to as a Barr body, and remains persistently in a transcriptionally silent state. X-chromosome inactivation exists in two different forms: random and imprinted, differing in terms of their developmental timing and mechanism of action besides using the same RNAs and silencing enzymes (Ahn & Lee, 2018).
Embryology and Theory of Evolution
Embryology plays a vital role in understanding biological evolution and facilitates the identification of similarities and distinctions among different species. An embryo, originating from a fertilized egg, undergoes species-specific developmental processes. Observing the progression from embryo to adult in animals allows for comparisons with other organisms, aiding in the identification of evolutionary parallels. While early-stage embryos often exhibit similar appearances across species, divergences become more apparent as development proceeds. Organisms with closer genetic bonds tend to share a more prolonged period of similar embryonic development due to a more recent common ancestor. Consequently, embryology serves as compelling evidence supporting the theory of evolution and the divergence of species from a shared ancestry (Gilbert, 2014).
Charles Darwin's theory of evolution significantly influenced comparative embryology by highlighting embryonic resemblances as compelling evidence for the genetic interconnectedness of different animal groups. Darwin observed that embryonic organisms occasionally develop structures inappropriate for their adult forms (such as eyes in embryonic moles, pelvic rudiments in embryonic snakes, and teeth in embryonic baleen whale) but indicative of their relatedness to other animals. He argued that adaptations allowing organisms to thrive in specific environments tend to emerge late in the embryo's development. Darwin's recognition of "descent with modification" introduced two perspectives: one emphasizing common descent by highlighting embryonic similarities between animal groups and another emphasizing modifications by showcasing how development changes to produce structures facilitating adaptation to specific conditions. This dual perspective enriched the understanding of evolutionary processes through embryological evidence (Gilbert, 2014).

Phenotypic plasticity is characterized by the capacity of individual genotypes to generate diverse phenotypes in response to varying environmental conditions (Pigliucci et al., 2006). This encompasses the capability to adjust developmental paths based on specific environmental cues. Moreover, phenotypic plasticity involves an organism's ability to alter its phenotypic state or activities, such as metabolic processes, in response to fluctuations in environmental conditions, enabling organisms to adapt to environmental unpredictability and heterogeneity While there is a widespread acknowledgment of the significance of phenotypic plasticity as a crucial attribute of developmental systems, its role in adaptive evolution remains a topic of debate. For example, the consensus acknowledges that phenotypic plasticity can enhance organism survival under specific conditions, however, there is no agreement on whether plasticity has the potential to instigate the evolution of novel traits and foster taxonomic diversity (Fusco & Minelli, 2010). The discussion also remains open regarding whether phenotypic plasticity predominantly accelerates or retards the course of evolutionary change.
As mentioned above, evolutionary alterations in the genome have the potential to induce changes in development, a phenomenon that follows the idea of natural selection. This intertwining of evolutionary and developmental processes introduces a dynamic where the outcomes of development impose constraints on genome evolution. To be more specific, natural selection acting on the results of developmental processes influences the evolution of genomic elements that play active roles in development. Presently, different prevalent models seek to establish a connection between developmental and evolutionary biology by leveraging morphological similarities observed during the developmental stages (Liu & Robinson-Rechavi, 2018).
Genetics and Evo-Devo
In genetics, homology between protein or DNA sequences is established based on shared ancestry, arising from either speciation events (orthologs) or duplication events (paralogs). A trait recognized as homologous is often termed a homolog, applicable to both a homologous protein and the encoding gene (DNA sequence). Partial homology is a concept where only a portion of the sequences compared is presumed to share common ancestry, while the remaining fraction does not.

Homologous sequences are classified as orthologous if they stem from a common ancestral gene and were separated by a speciation event. In simpler terms, when a species diverges into two separate ones, the copies of a single gene in these species are considered orthologous. These orthologs, or orthologous genes, exhibit significant sequence similarity and shared functional domains, indicating their common origin from a single gene in the last common ancestor. Understanding orthologs helps researchers trace the evolutionary relationships and developmental patterns across different species. On the other hand, homologous sequences are paralogous if they arise from a gene duplication event within the same genome. This duplication provides opportunities for genetic divergence, leading to the evolution of new functions (Brigandt, 2007).
In Evo-Devo, the study of paralogs contributes to understanding how changes in gene copies within a species can impact development and contribute to evolutionary innovation. Paralogous genes can belong to the same species but may also exist in different species. For instance, the hemoglobin gene in humans and the myoglobin gene in chimpanzees are considered paralogs. Paralogs can further be categorized into in-paralogs (arising after a speciation event) and out-paralogs (arising before a speciation event) (6.3C: Homologs, Orthologs, and Paralogs, 2023).
Evo-Devo and Homology
Evolutionary developmental biology aims to understand how the processes of development have evolved over time, contributing to changes in the shape and size of organisms. So, it involves comparing the development of entire bodies or their subunits within the same organism or between organisms that are distantly related. Some developmental biologists, as part of evo-devo, delve into understanding how changes in gene expression and function influence alterations in body shape and patterns. This approach represents a shift in the common framework of traditional biological disciplines such as comparative morphology which sought homological structures or "sameness" across different organisms.

The incorporation of evo-devo principles has influenced and transformed the traditional understanding of biological disciplines by prompting a reevaluation of concepts like homology, where similarities in structures were considered evidence of shared ancestry. Instead, evo-devo emphasizes the dynamic relationship between development and evolution, shedding light on how genetic changes contribute to morphological variations. Features of organisms' body plans, known as bauplan features, become more stabilized during the evolution of modular organisms like vascular plants, arthropods, and vertebrates. These features are understood, to some extent, as adaptations to the environment (Rutishauser & Moline, 2005).
Evo-devo, with its integrative approach, has broadened the comprehension of the links between development and evolution, providing a richer understanding of the diversity and adaptations observed in living organisms. Scientists now know that despite undergoing over 1 billion years of independent evolution in various lineages, numerous animal toolkit proteins frequently demonstrate functionally equivalent activities in vivo when they are interchanged. These findings suggest that the biochemical characteristics of these proteins, along with their interactions with receptors, cofactors, and other molecular components, have undergone minimal divergence over extended periods of time. One well-known illustration of this functional interchangeability is observed in the ability of the mouse Pax-6 protein to induce the formation of ommatidia (simple eyes) in Drosophila, akin to the function performed by the Drosophila Pax-6 (Eyeless) protein. Another example would be the capacity of a cnidarian Achaete-Scute homolog (CnASH) to stimulate the formation of sensory organs in Drosophila, similar to the role played by the Drosophila homologs. This discovery underscores the surprising conservation of function across species, despite over one billion years of independent evolutionary divergence (Carroll, 2008).
Conclusion
The intertwined fields of evolutionary developmental biology, genetics, and embryology provide a comprehensive framework for understanding the processes underlying biological evolution. The close examination of genetic foundations, alterations in DNA, and the dynamic interplay between development and evolution elucidate the mechanisms driving morphological diversity. From the discovery of master regulatory genes like Hox genes to the exploration of epigenetic processes, these interdisciplinary approaches offer profound insights into the conservation and divergence of biological structures. Additionally, the reinterpretation of homology through an evo-devo lens emphasizes the importance of understanding not just shared structures but the dynamic processes that govern their evolution. The integration of evolutionary developmental biology, genetics, and embryology fosters a comprehension of the tapestry of life, unveiling the link between genetic inheritance and developmental pathways that shape the diverse array of organisms that can be observed in the natural world.
Bibliographical References
Agrawal, N., Dasaradhi, P. V., Mohmmed, A., Malhotra, P., Bhatnagar, R. K., & Mukherjee, S. K. (2003). RNA interference: biology, mechanism, and applications. Microbiology and molecular biology reviews : MMBR, 67(4), 657–685. https://doi.org/10.1128/MMBR.67.4.657-685.2003
Ahn, J., & Lee, J. (2008). X Chromosome Inactivation | Learn Science at Scitable. Www.nature.com. https://www.nature.com/scitable/topicpage/x-chromosome-x-inactivation-323/
Al, N. M., Simpson, B., & Ishwarlal Jialal. (2019, April 5). Genetics, Epigenetic Mechanism. Nih.gov; StatPearls Publishing. https://www.ncbi.nlm.nih.gov/books/NBK532999/
Bannister, A. J., & Kouzarides, T. (2011). Regulation of chromatin by histone modifications. Cell research, 21(3), 381–395. https://doi.org/10.1038/cr.2011.22
Brigandt, I. (2007). Typology now: homology and developmental constraints explain evolvability. Biology & Philosophy, 22(5), 709–725. https://doi.org/10.1007/s10539-007-9089-3
Carroll, S. B. (2008). Evo-Devo and an Expanding Evolutionary Synthesis: A Genetic Theory of Morphological Evolution. Cell, 134(1), 25–36. https://doi.org/10.1016/j.cell.2008.06.030
Chan, S. S., & Kyba, M. (2013). What is a Master Regulator?. Journal of stem cell research & therapy, 3, 114. https://doi.org/10.4172/2157-7633.1000e114
Duverger, O., & Morasso, M. I. (2008). Role of homeobox genes in the patterning, specification, and differentiation of ectodermal appendages in mammals. Journal of cellular physiology, 216(2), 337–346. https://doi.org/10.1002/jcp.21491
Elhamamsy A. R. (2017). Role of DNA methylation in imprinting disorders: an updated review. Journal of assisted reproduction and genetics, 34(5), 549–562. https://doi.org/10.1007/s10815-017-0895-5
Fedoriw, A., Mugford, J., & Magnuson, T. (2012). Genomic imprinting and epigenetic control of development. Cold Spring Harbor perspectives in biology, 4(7), a008136. https://doi.org/10.1101/cshperspect.a008136
Fusco, G., & Minelli, A. (2010). Phenotypic plasticity in development and evolution: facts and concepts. Introduction. Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 365(1540), 547–556. https://doi.org/10.1098/rstb.2009.0267
Gilbert, S. F. (2014). Evolutionary Embryology. Nih.gov; Sinauer Associates. https://www.ncbi.nlm.nih.gov/books/NBK10049/
Hall, B.K. (1999). Evolution and Development: Terms and Concepts. In: Evolutionary Developmental Biology. Springer, Dordrecht. https://doi.org/10.1007/978-94-011-3961-8_1
Hall, B. K. (2012). Evolutionary Developmental Biology (Evo-Devo): Past, Present, and Future. Evolution: Education and Outreach, 5(2), 184–193. https://doi.org/10.1007/s12052-012-0418-x
Jambhekar, A., Dhall, A., & Shi, Y. (2019). Roles and regulation of histone methylation in animal development. Nature Reviews Molecular Cell Biology, 20(10), 625–641. https://doi.org/10.1038/s41580-019-0151-1
Kitazawa, R., Haraguchi, R., & Kitazawa, S. (2023). Histone Modification in Histochemistry and Cytochemistry. Acta histochemica et cytochemica, 56(3), 41–47. https://doi.org/10.1267/ahc.23-00014
Kouzarides, T. (2007). Chromatin Modifications and Their Function. Cell, 128(4), 693–705. https://doi.org/10.1016/j.cell.2007.02.005
Liu, J., & Robinson-Rechavi, M. (2018). Developmental Constraints on Genome Evolution in Four Bilaterian Model Species. Genome biology and evolution, 10(9), 2266–2277. https://doi.org/10.1093/gbe/evy177
Loscalzo, J., & Handy, D. E. (2014). Epigenetic modifications: basic mechanisms and role in cardiovascular disease (2013 Grover Conference series). Pulmonary circulation, 4(2), 169–174. https://doi.org/10.1086/675979
Melnyk, C. W., Molnar, A., & Baulcombe, D. C. (2011). Intercellular and systemic movement of RNA silencing signals. The EMBO Journal, 30(17), 3553–3563. https://doi.org/10.1038/emboj.2011.274
Moosavi, A., & Motevalizadeh Ardekani, A. (2016). Role of Epigenetics in Biology and Human Diseases. Iranian biomedical journal, 20(5), 246–258. https://doi.org/10.22045/ibj.2016.01
Muhammad, T., Zhang, F., Zhang, Y., & Liang, Y. (2019). RNA Interference: A Natural Immune System of Plants to Counteract Biotic Stressors. Cells, 8(1), 38. https://doi.org/10.3390/cells8010038
NCBI. (2017). RNA Interference (RNAi). Nih.gov. https://www.ncbi.nlm.nih.gov/probe/docs/techrnai/
Paulsen, M., & Ferguson-Smith, A. C. (2001). DNA methylation in genomic imprinting, development, and disease. The Journal of Pathology, 195(1), 97–110. https://doi.org/10.1002/path.890
Rutishauser, R., & Moline, P. (2005). Evo-devo and the search for homology (“sameness”) in biological systems. Theory in Biosciences, 124(2), 213–241. https://doi.org/10.1007/bf02814485
Shahid, Z., Simpson, B., Miao, K. H., & Singh, G. (2024). Genetics, Histone Code. PubMed; StatPearls Publishing. https://pubmed.ncbi.nlm.nih.gov/30860712/#:~:text=The%20%E2%80%9Chistone%20code%E2%80%9D%20is%20a
Weinhold, B. (2006). Epigenetics: the science of change. Environmental health perspectives, 114(3), A160–A167. https://doi.org/10.1289/ehp.114-a160
Wolpert, L. (2000). What is evolutionary developmental biology? Novartis Foundation Symposium, 228, 1–14; discussion 46-52. https://doi.org/10.1002/0470846631.ch1
6.3C: Homologs, Orthologs, and Paralogs. (2023, March 15). Biology LibreTexts. https://bio.libretexts.org/Courses/Universiti_Putra_Malaysia/ESC4103_-_Environmental_Microbiology_(Universiti_Putra_Malaysia)/06%3A_Microbial_biotechnology/6.03%3A_Bioinformatics/6.3C%3A_Homologs_Orthologs_and_Paralogs
Visual Sources
Figure 1: Cheatle Jarvela, A. M., & Pick, L. (2016, January 1). Chapter Fourteen - Evo-Devo: Discovery of Diverse Mechanisms Regulating Development (P. M. Wassarman, Ed.). ScienceDirect; Academic Press. https://www.sciencedirect.com/science/article/abs/pii/S0070215315001805
Figure 2: Hall, B. K. (2003). Unlocking the Black Box between Genotype and Phenotype: Cell Condensations as Morphogenetic (modular) Units. Biology & Philosophy, 18(2), 219–247. https://doi.org/10.1023/a:1023984018531
Figure 3: Mark, M., Rijli, F. M., & Chambon, P. (1997). Homeobox Genes in Embryogenesis and Pathogenesis. Pediatric Research, 42(4), 421–429. https://doi.org/10.1203/00006450-199710000-00001
Figure 4: Chromatin structure, including histones and DNA. antibodies-online.com. From: https://www.antibodies-online.com/areas/epigenetics/
Figure 5: DNA Methylation | EpiGentek. (n.d.). Www.epigentek.com. https://www.epigentek.com/catalog/dna-methylation-c-75_21.html
Figure 6: Themes, U. F. O. (2016, July 3). Epigenetics. Musculoskeletal Key. https://musculoskeletalkey.com/epigenetics/
Figure 7: Chromatic remodelling. Nordic Biosite. From:https://nordicbiosite.com/news/chromatin-remodeling-and-unraveling-the-histone-code
Figure 8: Coco, C., Sgarra, L., Potenza, M. A., Nacci, C., Pasculli, B., Barbano, R., Parrella, P., & Montagnani, M. (2019). Can Epigenetics of Endothelial Dysfunction Represent the Key to Precision Medicine in Type 2 Diabetes Mellitus?. International journal of molecular sciences, 20(12), 2949. https://doi.org/10.3390/ijms20122949
Figure 9: Linz, D. M., Hu, Y., & Moczek, A. P. (2020). From descent with modification to the origins of novelty. Zoology, 143, 125836. https://doi.org/10.1016/j.zool.2020.125836.
Figure 10: Homology Terminology: Never Say the Wrong Word Again. (2016, July 9). Bitesizebio.com. https://bitesizebio.com/26762/homology-terminology-never-say-wrong-word/
Figure 11: EvoDevo. EvoDevo Universe. From: https://evodevouniverse.com/wiki/Evolutionary_development_%28evo_devo,_ED%29


Comments